


 Toll Free 1800 572 9282
Toll Free 1800 572 9282  mailus@wbcsmadeeasy.in
mailus@wbcsmadeeasy.in




















































































Botany Notes – Evolution Of Sporophytes – For W.B.C.S. Examination.
উদ্ভিদ নোট – স্পোরোফাইটগুলির বিবর্তন – WBCS পরীক্ষা।
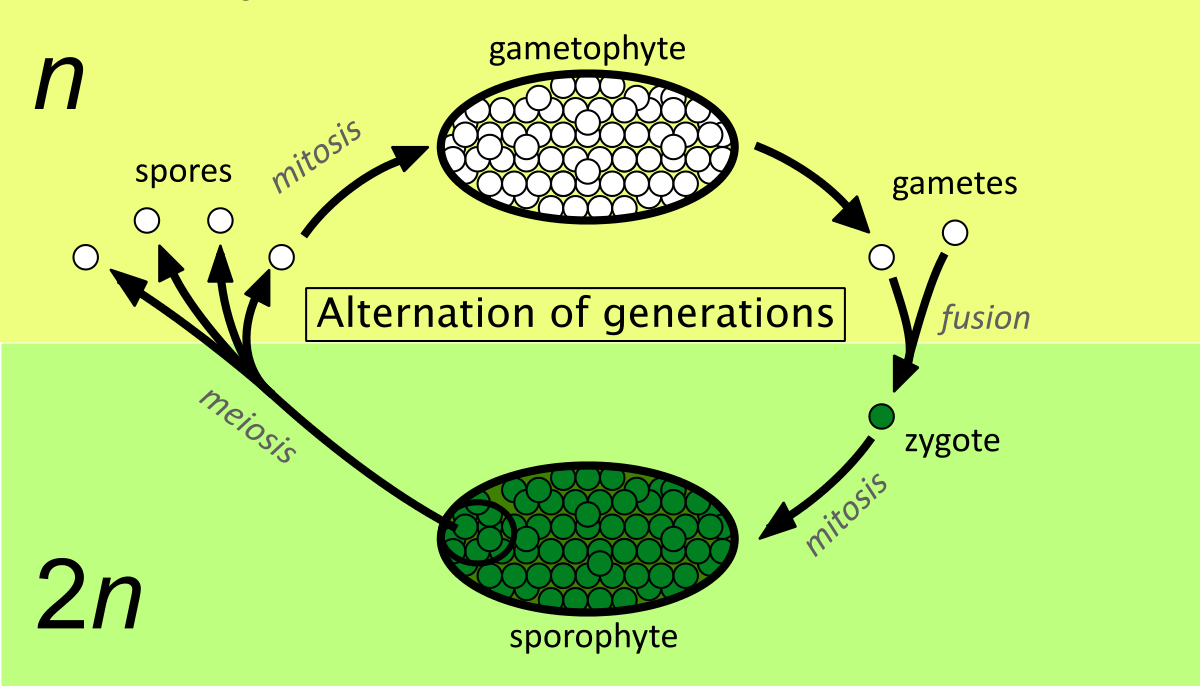
The sporophyte of bryophytes is called sporogonium which generally consists of a single, terminal sporangium (monosporangiate) with a bulbous foot and with or without an unbranched stalk or seta. The sporogonium is very delicate, short-lived and nutritionally dependent on its gametophyte.The sporophytic phase begins with the formation of a diploid zygote within the venter of the archegonium. In the simplest form of sporophyte (e.g., Riccia) the entire zygote is taking part in the formation of stelile capsule wall and the central sporogenous cells. In complex forms, zygote differentiates and sporogenous cells form more sterile tissues.This theory was advocated by Bower (1908- 35) and supported by Cavers (1910) and Campbell (1940). According to this theory, the primitive sporophyte of bryophytes was simple and most of the sporogenous tissue was fertile (e.g., Riccia) and from such a sporophyte, the more complex sporophytes (e.g., mosses) have been evolved by the progressive sterilisation of potential sporogenous tissue. This theory is also known as “theory of sterilisation”.Continue Reading Botany Notes – Evolution Of Sporophytes – For W.B.C.S. Examination.
The simple sporophyte of Riccia consists of a single-layered sterile jacket enclosing sporogenous cells with a very few absorptive nutritive cells (nurse cells). The zygote divides by a transverse wall, followed* by a vertical wall to form a four-celled embryo. Subsequently 20-30 celled embryo is formed by further divisions, in which periclinal divisions differentiate a single layered outer amphithecium and the inner multicellular mass, the endothecium.
Here the zygote has no polarity. The amphithecium forms the sterile jacket while the whole sporogenous cells (endothecium) differentiates into spores with a very few sterile nurse cells, possibly the forerunners of elaters.
Second stage:
In this stage, the zygote divides transversely to form a hypobasal and an epibasal cells. A small foot is formed from the hypobasal cell. The epibasal cells differentiates into an outer amphithecium and inner endothecium.
The amphithecium forms a single-layered sterile jacket of the capsule, while the endothecium differentiates into fertile sporocytes and long sterile elater-like nurse cells without the thickening bands. Thus, the zygote has polarity showing more sterilisation of sporogenous cells like nurse cells and sterile foot. This stage has been noted in Corsinia.
Third stage:
The development of sporophyte is like that of Corsinia, but there is more sterilisation of sporogenous tissue. This condition is noted in Sphaerocarpus sporophyte which consists of a sterile bulbous foot, a narrow sterile seta developed from hypobasal cell and a fertile capsule developed from endothecium containing sporocytes and sterile nurse cells.
Fourth stage:
This stage is represented by Targionia, where the sporophyte consists of a sterile bulbous foot, a sterile narrow seta and a fertile capsule. Here about half of the endothecial cells produce fertile sporogenous tissue, while the remaining half gives rise to sterile elaters with 2-3 spiral thickening. Hence, in Targionia, more sterilisation of sporogenous tissue has been observed.
Fifth stage:
This stage is illustrated by Marchantia, where further sterilisation of sporogenous tissue has been noted in comparison with Targionia. In Marchantia, the sterile tissue consists of a broad foot, a massive seta, a single-layered jacket of capsule, sterile apical cap at the apex of capsule and a large number of long elaters with spiral thickening.
Sixth stage:
This stage is represented by some members of Jungermanniales like Pellia, Riccarclia, etc. Here more sterilisation of sporogenous tissue has been observed. Sporophyte is differentiated into foot, seta and capsule having multilayered jacket. The sporogenous tissues produce mass of sterile elatophores and diffuse elaters.
Seventh stage:
This stage is illustrated by members of Anthocerotophyta like Anthoceros. Here marked reduction in the sporogenous tissue has been noted. The multilayered capsule differentiates into epidermis with stomata and chlorophyllous cells.
The central columella derived from endothecium is composed of 16 vertical rows of sterile cells. The further sterilisation of sporogenous tissue has been observed in the formation of pseudoelaters which are elongated 3-4 celled, simple or branched structure without thickening band.
Eighth stage (Final stage):
The members of Bryopsida like Funaria, Polytrichum, Pogonatum etc., show the highest degree of sterilisation. The sporophyte is differentiated into a foot, a long seta and a capsule. The sterile tissue of capsule consists of the apophysis, operculum, many- layered jacket, the columella, trabeculae, the wall of spore sac and the peristome. The sporogenous tissue is restricted to the spore sacs only, hence it forms a negligible portion in the sporophyte.
(ii) Theory of Regressive evolution i.e., evolution of sporophytes due to the progressive reduction or simplification:
This theory is known as regressive or retrogressive theory, and supported by several scientists like Church (1919), Kashyap (1919), Goebel (1930) and Evans- (19391 According to this theory, the most simple sporophyte of Riccia (comprised of a simple capsule) is the most advanced type which has been evolved by the simplification or progressive reduction of the complex sporophytes (foliose with complex assimilatory tissue and functional stomata) of mosses (e.g. Funaria, Pogonatum, Polytrichum etc.).
The stages of progressive reduction of the foliose sporophyte (primitive type) to the simpler sporophyte (advanced type) have been enumerated:
(a) The semiparasitic foliose sporophyte gradually lost its leaves and became embedded within the gametophyte.
(b) There is a gradual reduction of the assimilatory (photosynthetic) tissue in the sporophytes and subsequently this tissue is confined only to the jacket of capsule (e.g., Funaria, Anthoceros).
(c) Stomata are restricted in the apophysis region (e.g. Funaria, Polytrichum) that communicate with the intercellular spaces. In Sphagnum, the stomata of apophysis are non-functional and become rudimentary. In all liverwort members stomata are completely absent in sporophytes.
(d) The capsules of most mosses (Funaria, Polytrichum, Sphagnum, etc.), hornwort (Anthoceros) and some jungermanniales (Pellla, Porella) are multilayered which subsequently became single-layered (Marchantia, Plagiochas- ma, Riccia) by reduction.
(e) The foot and seta are well-developed in mosses (Pogonatum, Funaria, etc.) and some liverworts (Pellia, Marchantia, etc.). The seta became much reduced and form a narrow sterile part of the sporophyte (Corsinia, Targionia).
In hornworts, the sporophyte is made up of a foot and an elongated capsule only, seta is absent. Finally, in Riccia foot and seta are absent and the sporophyte is represented by a single capsule only, which is supposed to be the most simple as well as advanced sporophyte among bryophytes.
(f) The sporophytes of mosses show the highest degree of sterilisation with a negligible amount of sporogenous tissue. There has been gradual reduction in the sterile tissue of the capsule, with simultaneous increase in the amount of sporogenous tissue.
In hornworts, a good amount of sporogenous tissue is formed from the inner layer of amphithecium. In liverworts (Riccia, Marchantia) the entire endothecium gives rise to sporogenous cells.
Evolution of Gametophytes in Bryophytes:
The evolution of thalli in bryophytes is a much disputed problem. There is no substantial fossil evidences of bryophytes that support to the sequential evolution theory of gametophytes among bryophytes.
There are two opposing theories regarding the nature of the vegetative structure of the primitive bryophytic gametophyte and its subsequent evolution:
1. The upgrade or the progressive evolution theory, and
2. The downgrade or the regressive evolution theory.
1. The upgrade or the progressive evolution theory:
According to this theory, the primitive gametophyte was a simple, dorsiventral, prostrate thallus, both in external as well as in internal forms. Cavers (1910) and Campbell (1891-1940) were the main proponents of this theory. The evolution of gametophytes took place from liverworts to mosses in an ascending series of gradually increasing complexity with regard to the organisation of internal tissue and sex organs.
According to Cavers the ancestor gametophyte resembles the present day Sphaerocarpus and Marchantiales has been considered as a blind line of evolution from the hypothetical Sphaero- Riccia. While Campbell suggested that thalli of the present day Riccardia and Metzgeria resemble the simplest ancestral gametophyte. From the simple thallus, the evolution of complex gametophytes took place in two different lines.
In the first line, the gametophyte retained its external simple, thallose form as found in Marchantiales. Simultaneously there was a gradual increase in complexity in cellular organisations. This has been evidenced by the nature of pores, air chambers and the aggregation of sex organs in a definite receptacle (e.g. Marchantia).
The sexual receptacles show a wide range of organisation. In Riccia, the individual sex organs are scattered over the median portion of the thallus. In Marchantia, the sex organs are borne on a complex stalked receptacle called gametophore.
An intermediate condition in between the Riccia and Marchantia has also been observed where sex organs are aggregated into a cushion-like or ridge-like receptacle. These receptacles are borne on the thallus — dorsally or terminally.
In the second line, the gametophytes retained their simple internal structure (lack of airpores and air chambers). But there was a gradual elaboration of the external part of the gametophyte leading to the formation of the appendicular organs.
2. The downgrade or the regressive evolution theory:
According to the downgrade theory, the primitive gametophyte was an erect leafy shoot having radial symmetry (members of Calobryales and true mosses). From such ancestral forms the dorsiventral thalli of liverworts and hornworts got evolved in reverse direction i.e., regression of increasing simplicity.
Among the proponents of the downgrade theory, Wettstein (1903-1908), Church (1919), Evans (1939), Goebel (1930) and Kashyap (1919) are the most prominent scientists.
Kashyap (1919) advocated the regressive evolution from the results of his extensive studies of Indian Marchantiales.
The principal points in the reduction series from Marchantia as the basic type along the various phyletic lines have been summarised below:
(a) Reduction in the number of involucre:
In Marchantia, sex organs are well-protected by many involucres. A gradual reduction in the number of the involucres has been observed in Conocephallum, Aitchinsoniella and Exormo- theca, which has been finally culminating in Tarefionia with a single involucre .
(b) The loss of assimilatory filaments in the air chamber:
A gradual reduction series has been noted in many members (Fig. 6.64). In Marchantia and Preissia, the thallia show complexity in having air pores and air chambers full of assimilatory filaments. There is a gradual reduction in the assimilatory filaments in Conocephallum conicum (the filaments are short in the air chambers), Wiesnerella decundata (the filaments rudimented into papillate cells). The assimilatory filaments ultimately disappear in the aquatic Dumortiera hirsuta .
(c) Simplification of Pores:
In complex forms like Marchantia and Preissia, the pores are complex, barrel-shaped and present both on the thallus and the discs of the gametophores. In Conocephallum and Reboulia, discs bear only barrel-shaped pores, while thalli bear only simple pores.
In Exormotheca and Stephansoniella, the pores are simple both on the thallus and in the discs. The well-defined pores are totally absent in Riccia .
(d) The gradual shifting of the stalks of antheridiophores and archegoniophores from the terminal to dorsal position:
Mehra (1969) proposed the above hypothesis. In Marchantia, the antheridia and archegonia are borne terminally on the stalked gametophores. In Preissia qua- drata and Plagiochasma articulatum, the stalk is initially terminal, but becomes dorsal by the further growth of the thallus.
A further downward shifting of the stalk is observed in Corsinia and Boschia, where the female receptacle almost becomes sessile by the elimination of the stalk .
Our own publications are available at our webstore (click here).
For Guidance of WBCS (Exe.) Etc. Preliminary , Main Exam and Interview, Study Mat, Mock Test, Guided by WBCS Gr A Officers , Online and Classroom, Call 9674493673, or mail us at – mailus@wbcsmadeeasy.in
Visit our you tube channel WBCSMadeEasy™ You tube Channel
Please subscribe here to get all future updates on this post/page/category/website












