


 +919674493673
+919674493673  mailus@wbcsmadeeasy.in
mailus@wbcsmadeeasy.in








































































Insect Endocrine Glands – Zoology Notes – For W.B.C.S. Examination.
প্রাণিবিদ্য নোট – পোকার অন্ত: স্র্রাবী গ্রন্থি – WBCS পরীক্ষা।
Neuroendocrine control mechanisms are observed in all animals that possess a nervous system. Recent analyses of neuroendocrine functions in invertebrate model systems reveal a great degree of similarity between phyla as far apart as nematodes, arthropods, and chordates. Developmental studies that emphasize the comparison between different animal groups will help to shed light on questions regarding the evolutionary origin and possible homologies between neuroendocrine systems. This review intends to provide a brief overview of invertebrate neuroendocrine systems and to discuss aspects of their development that appear to be conserved between insects and vertebrates.Continue Reading Insect Endocrine Glands – Zoology Notes – For W.B.C.S. Examination.
Endocrine and neuroendocrine cells
Cells in multicellular animals communicate through signaling mechanisms that take place at direct intercellular contacts, or that involve signals released systemically into the extracellular space where they diffuse over large distances and are able to affect targets far removed from the signaling source. The first mechanism, communication of cells that are in direct contact, is developed to a state of high complexity in the nervous system. Here, a multitude of signals in the form of neurotransmitters chemically couple networks of neurons at specialized cell–cell contacts, the synapses. The second mechanism of cell–cell communication defines the endocrine system. It involves secreted signals, hormones that affect target cells in a less directed way, since all cells expressing receptors for a given hormone will react when that hormone is released. The endocrine system in bilaterian animals consists of multiple specialized cell populations, sometimes compacted into glands that are found in all parts of the body, and are derived from all three germ layers (Tombes 1970, Highnam & Hill 1977, Gorbman et al. 1983, Laufer & Downer 1988). Endocrine glands regulate a large number of homeostatic mechanisms. They include the activity of neurons, muscles, and pigment cells during specific behaviors (food intake, fight and flight, and reproduction), the activity of visceral muscle and exocrine glands (digestion), the control of major metabolic pathways (synthesis, storage, and release of carbohydrates and lipids), the control of the ionic milieu through absorption and excretion, the formation and maturation of gametes, and growth and regeneration of the body. In many instances, endocrine glands form an integrated system in which hormonal production and release is controlled through feed back loops.
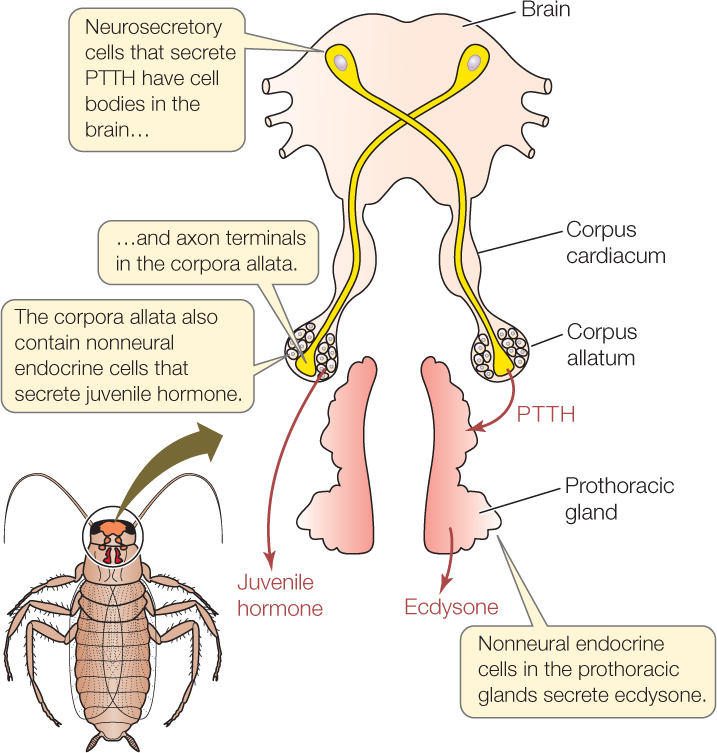
The neurosecretory system in insects consists of several sets of neurosecretory cells located in the brain and ventral nerve cord. The majority of NSCs are found in the dorso-medial protocerebrum, the so-called pars intercerebralis (PI) and pars lateralis (PL; Pipa 1978, Raabe 1989, Schooneveld 1998, Veelaert et al. 1998, Siegmund & Korge 2001). These NSCs project their axons toward a set of endocrine glands, the corpora cardiaca (CC) and corpora allata (CA; Fig. 3D). In Drosophila, the CC and CA, along with a third neuroendocrine gland, the prothoracic gland (PTG), are fused into a single complex, the ring gland, which surrounds the anterior tip of the dorsal blood vessel (Fig. 4D). Containing release sites for neurosecretory products, the CC and CA act as neurohemal organs. At the same time, neuropeptides that reach the CC and CA from the brain may act locally on the glandular cells of these organs and control the release of their hormones.
The pars intercerebralis comprises an unpaired cluster of neurons located along the anterior brain midline, flanked by the mushroom body on either side and the central complex ventrally. The architecture of the NSCs has been the object of many studies, describing them as monopolar neurons with dendrites spreading in the hemispheres and axons joining the first nerve to the corpora cardiaca (nccI; Geldiay & Edwards 1973, Rowell 1976, Koontz & Edwards 1980, Zaretsky & Loher 1983, Homberg et al. 1991a,b, Fig. 3D). NSCs of the PI secrete insulin-like peptides, FMRFamide-like peptides, Locusta-diuretic hormone, pigment-dispersing hormone, Manduca sexta-allatostatin, ovary ecdysteroidogenic hormone, and myomodulin (reviewed in Nassel 2002). The NSCs forming the PL of the brain produce FMRFamide-like peptides, pigment-dispersing hormone, corazonin, and M. sexta-allatostatin. Their axons form the second nerve of the corpora cardiaca (nccII), which in most insects travels alongside the nccI (Fig. 3D); in Drosophila, both nerves are enclosed by a single perineurial (glial) sheath.
Outside the PI and PL, the tritocerebrum and the ventral nerve cord, as well as the ganglia of the stomatogastric nervous system (SNS) contain neurosecretory cells. NSC axons of the tritocerebrum and subesophageal ganglion projecting toward the corpora cardiaca form the nccIII nerve (Penzlin 1985, Kim et al. 1998, Schooneveld 1998, Nassel 2002); neurosecretory axons from the SNS also form a compact axon bundle connecting the hypocerebral ganglion with the corpora cardiaca (Penzlin 1985). NSCs of the ventral nerve cord have release site associated with the dorsal glial sheath of the cord and the segmental peripheral nerves (Duve et al. 1988, Nassel et al. 1988, Schooneveld 1998).
Peripheral neuroendocrine glands in insects: the CC of insects consist of two distinct zones, an unpaired ventral storage lobe, containing the terminals of NSCs located in the PI and PL, and a more lateral glandular lobe that is formed by NSCs in its own right (Gupta 1990, Dai & Gilbert 1991, Dorn 1998, Schooneveld 1998). Some insects, among them flies, lack a storage lobe; here, neurosecretory axons that would terminate in the CC in locusts and other insects pass through the CC and end in contact with the aorta (King et al. 1966, Schooneveld 1998). The glandular lobe of the CC produces several hormones, including AKH, certain glycemic factors, cardiac-accelerating factors, and melanin-inducing factor. The AKH hormones are the major products of the CC, which are secreted into the hemolymph to mobilize lipids and carbohydrates during flight (O’Shea & Rayne 1992, Noyes et al. 1995, Veelaert et al. 1998, Nassel 1999, Oudejans et al. 1999).
The CA produces juvenile hormone (JH), a fatty acid derivative which has profound effects on larval growth, metamorphosis, egg development, and sexual behavior (Veelaert et al. 1998, Vullings et al. 1999, Gilbert et al. 2000). The pars lateralis in the brain via its projections to the CA is the source of positive and negative control over JH production (Stay et al. 1996, Stay 2000, Siga 2003). One of the neuropeptides reaching the CA was identified as allatostatin, which inhibits JH release.
The prothoracic gland (PTG) derives its name from the fact that in most insects it is situated in the prothoracic segments. In dipterans, the PTG consists of bilateral clusters of large glandular cells that form the lateral wings of the ring gland. The PTG synthesizes and secretes a polyhydroxylated steroid prehormone, which is then converted to the major molting hormone, 2O-hydroxyecdysone, by peripheral tissues (Bollenbacher et al. 1975, Gilbert et al. 1997). Ecdysone triggers the transition from larval to pupal molts. It is also responsible for the complex metamorphic-remodeling processes that shape the adult organs of the insect body. The level of ecdysone is controlled by numerous humoral and neural pathways. One of the factors that controls ecdysone release is prothoracicotropic hormone (PTTH), a peptide that has been isolated in several insect species, including Drosophila (Gilbert et al. 1997, Kim et al. 1997). Axon tracts that funnel PTTH (and other factors) directly to the PTG stem from the ventral nerve cord (prothorax!); in addition, PTTH-secreting NSCs are located in the PI, send their axons through the NCCI to the CA where they release PTTH into the hemolymph (Westbrook & Bollenbacher 1990, Dai et al. 1995, Aizono et al. 1997).\
Please subscribe here to get all future updates on this post/page/category/website












